
Abstract
In this paper, I explore the origins of narrative and the neuroscientific evidence in support of narratives. There is clear support that narrative construction and use are enabled by and emerge from our neurological architecture. This means that our narratives are contingent upon biology and evolutionary theory. Because narratives also facilitate complex socialization, the foundational narratives that serve as the keystone, cornerstone, or bedrock of any society also make all narratives that emerge out of those societies contingent upon those foundational, or sacred narratives. Together, the biological, evolutionary, and social spaces wherein we reside limit the stories that can structure our lives in a beneficial manner. This exploratory model highlights the notion that, with prudent and constrained use, narratives can improve the lives of people who write or read them.
Introduction
In the beginning was the Word, and the Word was with God, and the Word was God. -- John 1:1
Have you ever asked yourself why you should read, what purpose does it serve, why you seem to be enthralled by a certain kind of character or plot? I know I have. I have a deep fascination with the 2004 Battlestar Galactica series, Stargate SG-1, 30-Rock, and so on. We all have stories we enjoy and return to. But why do we enjoy them, what do we get out of them, why do we get enraptured by certain stories and not others? What might the effect of our narrative consumption be? To what extent might we alter our lives through narratives? In this paper, I hope to answer some of these questions by exploring a model for narrative and script construction via a neuroscientific framework.
I was inspired to write this paper and to explore this topic after listening to a podcast with Jordan Peterson and Angus Fletcher. I have also been doing research on Creativity and so, given that the two streams crossed one another, I thought it would be worthwhile to explore this kind of topic. The framework or model that I will be proposing in this paper is intended to help people better understand the stories they tell themselves and the stories we hear from others. This framework can also be used to structure the stories we tell ourselves, the narratives we tell ourselves, the narratives we encounter or experience, or other peoples’ narratives or tales to prepare us to avoid the pitfalls and conflicts that cause chaos or hell in our own lives. And lastly, I think this model may have the capacity to serve as a bedrock for the origin of narrative, or may even support any understanding of any evolutionary theory of narrative, as it is built upon a biological framework, which arose out of our phylogenetic history.
To start, I would like to define what I am talking about. Here’s the list of terms that will be defined: Phenomenon, narrator, narrative, script, scripture, paradigmatic structure, schema, scheme, scene, literature, and knowledge. To define these terms, I will be using the Chambers Dictionary of Etymology, which I will use to create working theoretical definitions for the model.
Definitions
The origin of the word Phenomenon is borrowed from Latin phaenomenon and Greek phainómenon, which means, that which appears or is seen, also, a fact, occurrence, or a manifestation. The important aspect of this word is that it not only provides us with a mental state for a person providing a concept or idea but also the manner by which that idea or concept was arrived at; through sight, or more broadly, through the senses. This is key as it restricts the derivation of ideas to that which can be observed, that which is part of our sensory experience; it does not permit the usage of ideas and concepts that lack substance, which are merely a matter of our own conception; I challenge the reader to come up with an idea that does not correspond to something from their experience or within their environment in some way if they believe this assertion is false. The importance of this facet of the model will be discussed later.
Secondly, the concepts Narrator, Narrative, and Narrate. Narrative has its origins in Latin, Middle French and English (Scottish Law), which come together to succulently define the term as, an account or recounting, via a story, that contains a statement or series of statements on the relevant or essential facts of an event, or phenomenon. To ensure that there are no loose ends in this definition, I will also have to define what an account is. The term Account is borrowed from Old French, and is thus a derivative of Latin meaning, a computation or calculation, which will be defined for our purposes as the act of calculating, which can be more thoroughly defined as the act of formulating, or figuring; giving an event structure, shape, or form through the use of language. Then Narrative can more completely be defined as an act of speech that gives an event structure, shape, or form, containing only relevant or essential facts about a phenomenon, or an event that was captured via the senses. Then the narrator is someone who gives such an account, and the act of providing a narrative can be called narrating, which stems from the infinitive, to narrate.
Next, Script and Scripture. When I was listening to the podcast with Jordan and Angus, I was going through some of these terms and a notion that caught me off guard was the idea that Literature and Scripture are synonymous. This paper does not hold this notion and will rather argue that Literature comes from Scripture and Narrative, and that neither are synonymous with the former. But before I explain why that is the case, Scripture.
To define Scripture, we must first define its root term, Script. The word Script has its origins in the Latin term scrïptum, which means a writing, a book, treatise, law, line, or mark. Scribes would have been the people who typically kept scripts, which has its roots in Latin, Greek, and Hebrew, which respectively are scrïba, grammateús, and söphër, which roughly translate as writer or scholar. In a stricter sense, in Latin, scrïba meant the keeper of accounts, or the narrator, and was derived from the Latin term for writing. In Hebrew the term söphër can also mean teacher, or a teacher of Jewish Law; a person who shows his students how to make proper marks, how to read and write for the sake of understanding Jewish Law. The term Scripture then has its roots in what is written, written characters, what’s composed, or inscribed by a teacher, scholar, or scribe, who is the person responsible for keeping account of the relevant facts of any given event, or providing a narrative for a phenomenon. The term Scripture seems to have only been given its connotation with the sacred writings of the Bible at a later date. Then in summation, Scripture is the collected scripts of scribes, who were teachers, scholars, or narrators for the phenomenon experienced by the Hebrews, Greeks, and Romans. These peoples are generally considered to have provided the foundational ideas of Western civilization, which is more than likely why their works are regarded as holy, sacred, or foundational; they are the keystone, cornerstone, or bedrock of Western Civilization.
Next, Schema, Scheme, and Scene. It is relevant to describe Scene first, which is borrowed from the Latin and Greek, scaena and skene, which respectively are defined as a stage, tent or booth. The Latin term for scene, meaning stage, can be defined via the Vulgar Latin, staticum, a place for standing, which should lead one to the word existence, or facts, if the reader is wise. The term Existence has its roots in the Latin terms existere, exsistere, stand forth, appear, exist, which if a scene is a place for standing, it is also a place where one may exist, be caused to stand; and because standing, existing is a deed, a scene is also a place where one may observe facts, which are events that have occurred, actions or deeds that have or do take place. Then scenes are places where a person may exist and where facts occur, which can be constrained by language via narrations, which can be recorded via a script or scripture. But how do we get a scene? A Schema.
A Schema is a shape, figure, or form; generally, an outline, design or diagram. Then Schemas are the collective shapes, figures, or forms we give scenes, likely with scripts, language, and thus our narrations. Schemes are the particular shapes, figures, and forms of the designs and diagrams of scribes, which comes from the Greek term schêma. This term is related to scheîn, to get, and échein, to have or hold. These Greek words are cognate with the Sanskrit terms sáhas, strength and victory; Indo-European seghos and in Germanic-Gothic sigis, victory; Old Icelandic, sigr, Old High German, sigu, Middle Dutch, sëghe, Old Frisian, sï, and Old English, sieg all meaning victory; from Proto-Germanic (Chambers Dictionary of Etymology, p 966). Clearly it seems as if a proper scheme will provide a person or people, success or victory, which comes from a proper schema or a series of figures or schemes that form a design or diagram, which produce a scene.
Because this idea may seem convoluted, I will try to describe this notion as succinctly as possible. Through the use of language, narrators and scribes produce narratives and scripts, which create a schema or diagram of proper action, which are made up of schemes or a series of figures, which provide a narrator or audience a scene which they can exist within, or observe a series of facts from, which ultimately provide a person with some type of success. Wouldn’t it be nice to have a term that encapsulates the scenes produced by this process and the facts and states of being within these scenes? Well that’s why we have the phrase, Paradigmatic Structure.
A Paradigm is defined as a pattern or example, which is borrowed from the Latin and Greek term paradïgma and parádeigma, from paradeiknýnai, which can be defined as a pattern, example, or that which is shown side by side, shown, pointed out. Then paradigmatic means of or like a paradigm, an example; that which serves as a pattern or example. Thus, Paradigmatic Structures are scenes that serve as examples and can be presented side-by-side for a reference, but to what? Reality. I will return to this point later in the paper.
Lastly, Literature and Knowledge. Literature is the meaning we take from a body of writings from a period or a country; and more loosely, the knowledge from books and book learning. This is why I think it is important to differentiate Scripture from Literature, and to recognize that they are not synonymous. Literature is what we get from Scripture or Narratives, and it may form a body of written documents (e.g., Neuroscientific Literature), but it is not scripture. Then what is Knowledge? Knowledge, has its roots in what is Known or that which can be recognized. The term Recognized may be defined as that which is identified, recalled to mind, examined and certified; that which is treated as valid. The term Valid comes from Middle French, valide, is borrowed from the Latin term Validus, which means effective or strong, from valëre, to be strong. In this sense, Knowledge is valid information that provides one with the truth, which affords one strength.
Once again, to succinctly convey the idea here: Literature is the meaning, a form of knowledge, we take from a body of writings from a period or a country through book learning, which provides us with knowledge. Knowledge is information that is recognized as valid, providing one with truth, enabling them to be strong. Then Scripture can provide us with Literature, which can provide us with Knowledge, which grants us strength through its validity. For those who have been following the argument, this would be why paradigmatic structures provide an individual (or group of people) with success (victory) if those paradigmatic structures relate to reality, are true, i.e., valid.
Model of the Paradigmatic Structure
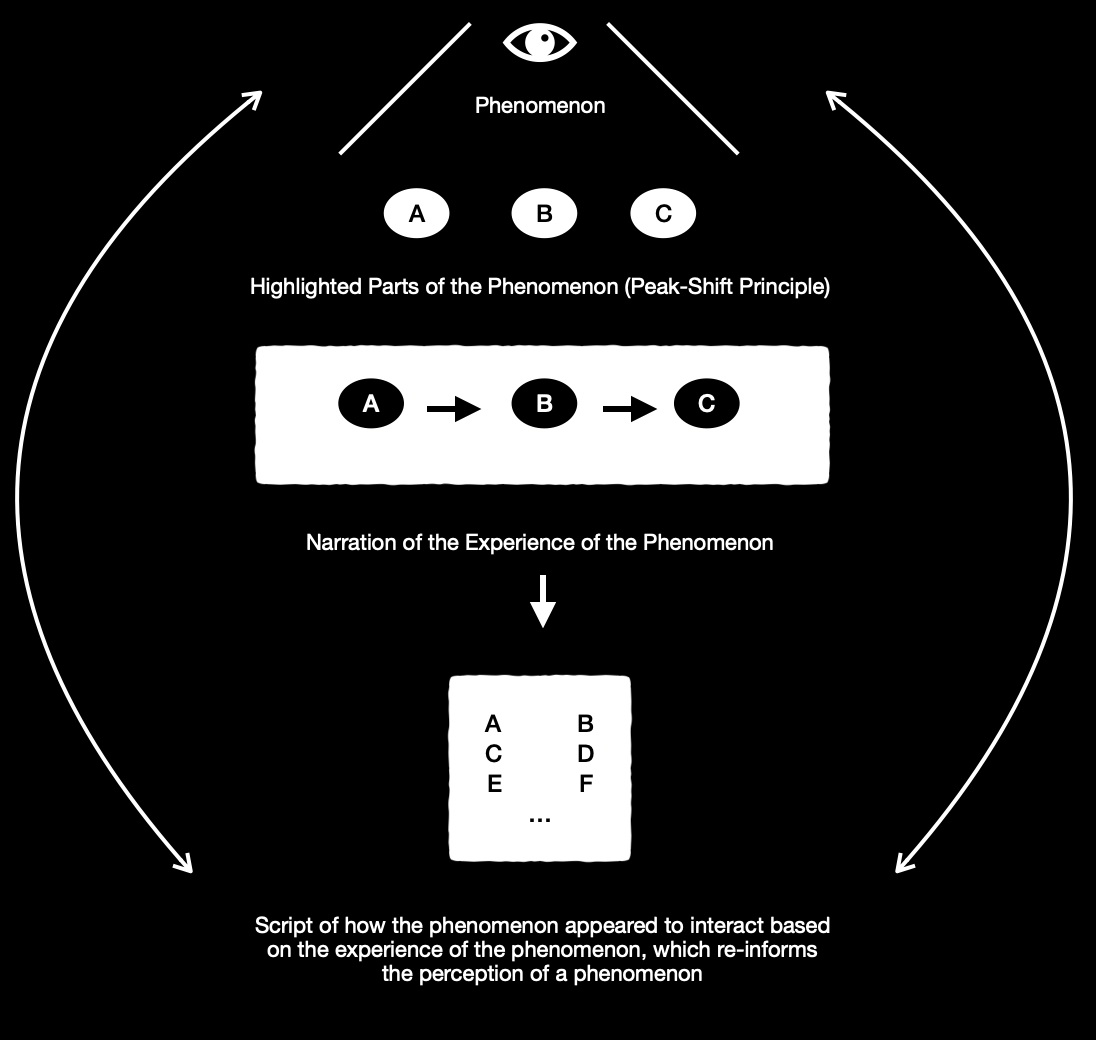
Fig. 1
In figure 1, the reader should be able to see an observer experiencing a phenomenon, the highlighted parts of that phenomenon, the narration of the experience of that phenomenon, and the script that emerges out of the narration of the experience of the phenomenon (how it appeared, how it behaved, what it was like, etc.). This script then re-informs the phenomenon and the stimulus or stimuli that served as the vehicles for the phenomenon. In neuroscience, when an observer highlights only the relevant parts of a phenomenon and then expounds on those aspects in a narrative, he is behaving in accordance with something known as the peak-shift principle (1.). This principle is fully fleshed out in a paper by Ramachandran and Herstein. Secondly, the re-information of stimuli, such that they give rise to new experiences, is also an aspect of cognition and can be manifested by the use of metaphor (2.). Overall, this process is oscillatory, but it also produces schemas.
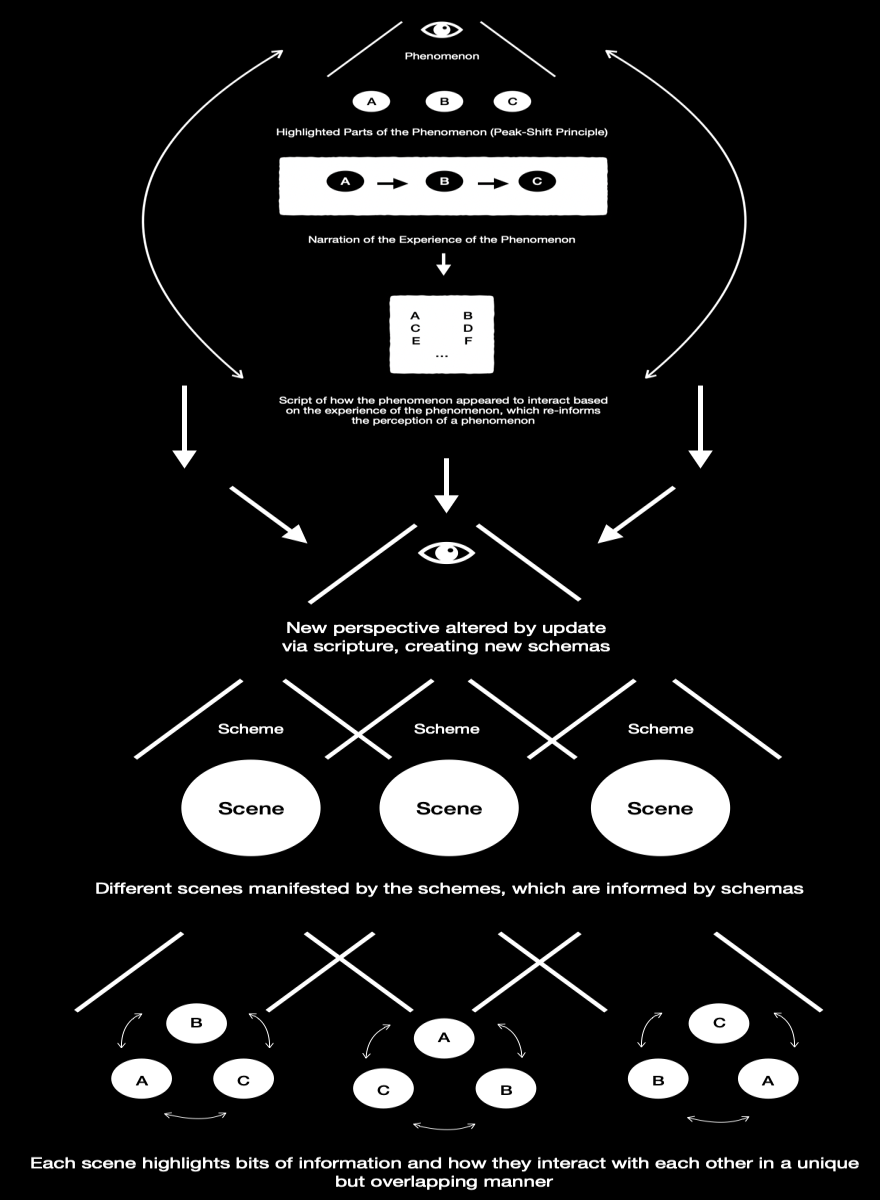
Fig. 2
Figure 2 expands on the processes that occur in the model overall, which stem from Figure 1. Figure 2 is differentiated by the fact that the script alters the perspective of the observer and narrator, who then creates either material or mental schema of his experiences, which can be particularized as schemes, which then produce different scenes. Each scheme, being different from the next, produces a different scene or perspective of the stimuli and thus the experience of the stimuli. These different scenes alter the information that is highlighted and how one experiences the parts of the stimuli. This does not mean that they are different bits of information, only that each scheme and thus scene, and perhaps even schema, alters how an observer experiences any of the parts of the stimuli; i.e., there are only the stimuli, but the way in which the stimuli is experienced is informed by the framework that’s used to observe them and the mental scene created by that framework. Examples of this phenomenon can be observed in, at least, both expert musicians and writers, who engage with their mediums differently than amateurs do (3. & 4.), and how training in music and writing can, perhaps, alter the perceptions of novices (5. & 6.).
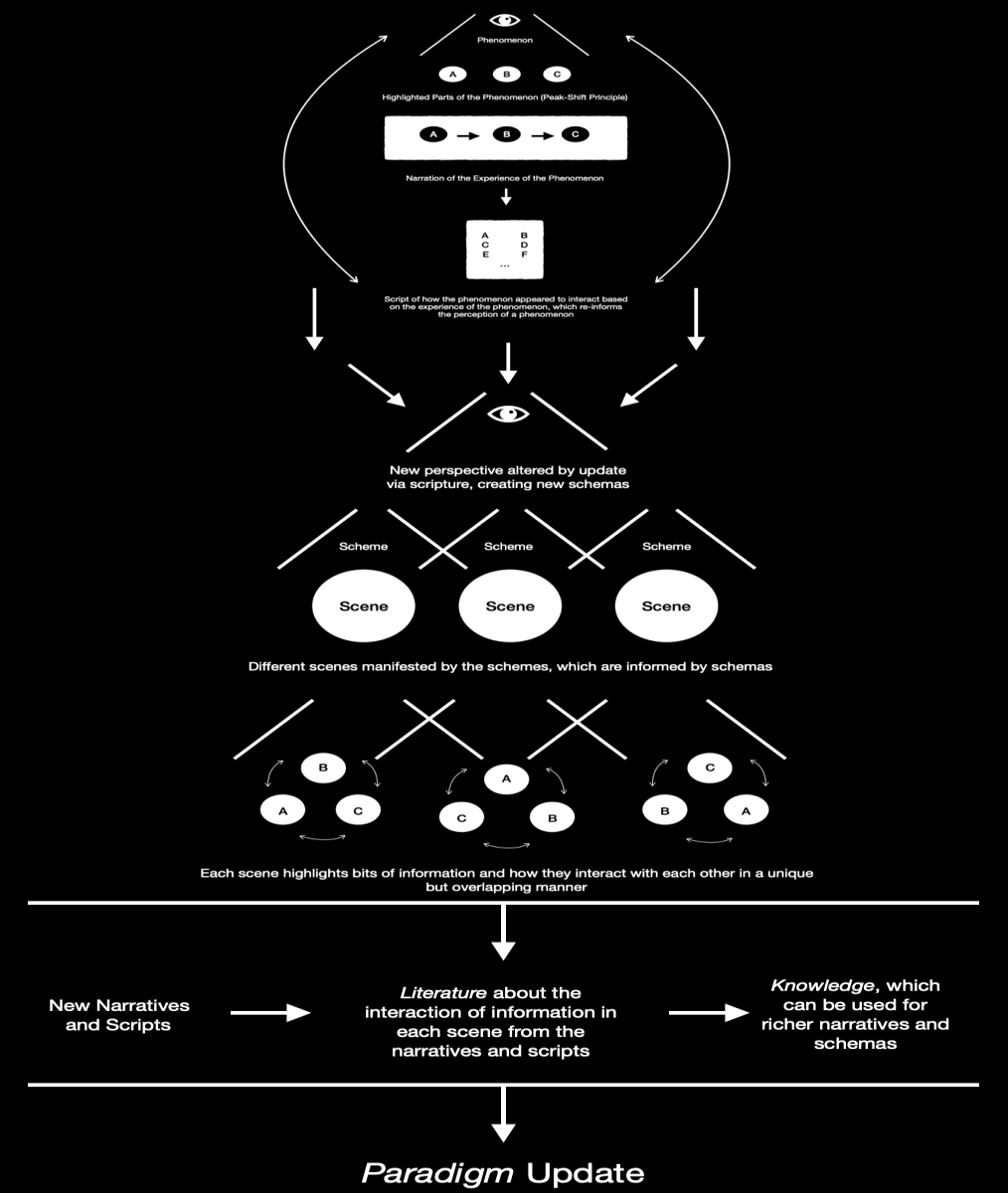
Fig. 3
Figure 3 expands on both Figure 1 and Figure 2 insofar as it highlights the fact that the processes of Figure 2 enable new narratives and scripts to be formed, which produces literature about those narratives and scripts, which finally enables an individual to gain access to knowledge about the stimuli and thus the world. The information is obviously informed by what the observer thinks, but given that it is knowledge and can be used to make predictions, the knowledge can either be regarded as valid or not, and thus capable of making the observer stronger, or not, which is the actual nature of truth. This process can be seen throughout history from the development of measures of creativity to the development of measures of intelligence (7. & 8.). In fact, throughout most academic fields, this continual process of script production, reproduction, development, and application can be observed. And of course, as any scriptwriter, novelist, or director can tell you, the script is never really finished, people just stop working on it (9.). The acquisition of knowledge in this manner, true knowledge, which may be differentiated from false knowledge based on its capacity to make accurate predictions about reality, results in a paradigm update, or a restructuring of our paradigmatic structures. Given that such structures serve as vehicles for our understanding of reality but are not reality, to have a sufficient update of one must result in something like a revelatory experience; a complete restructuring of how we experience reality.
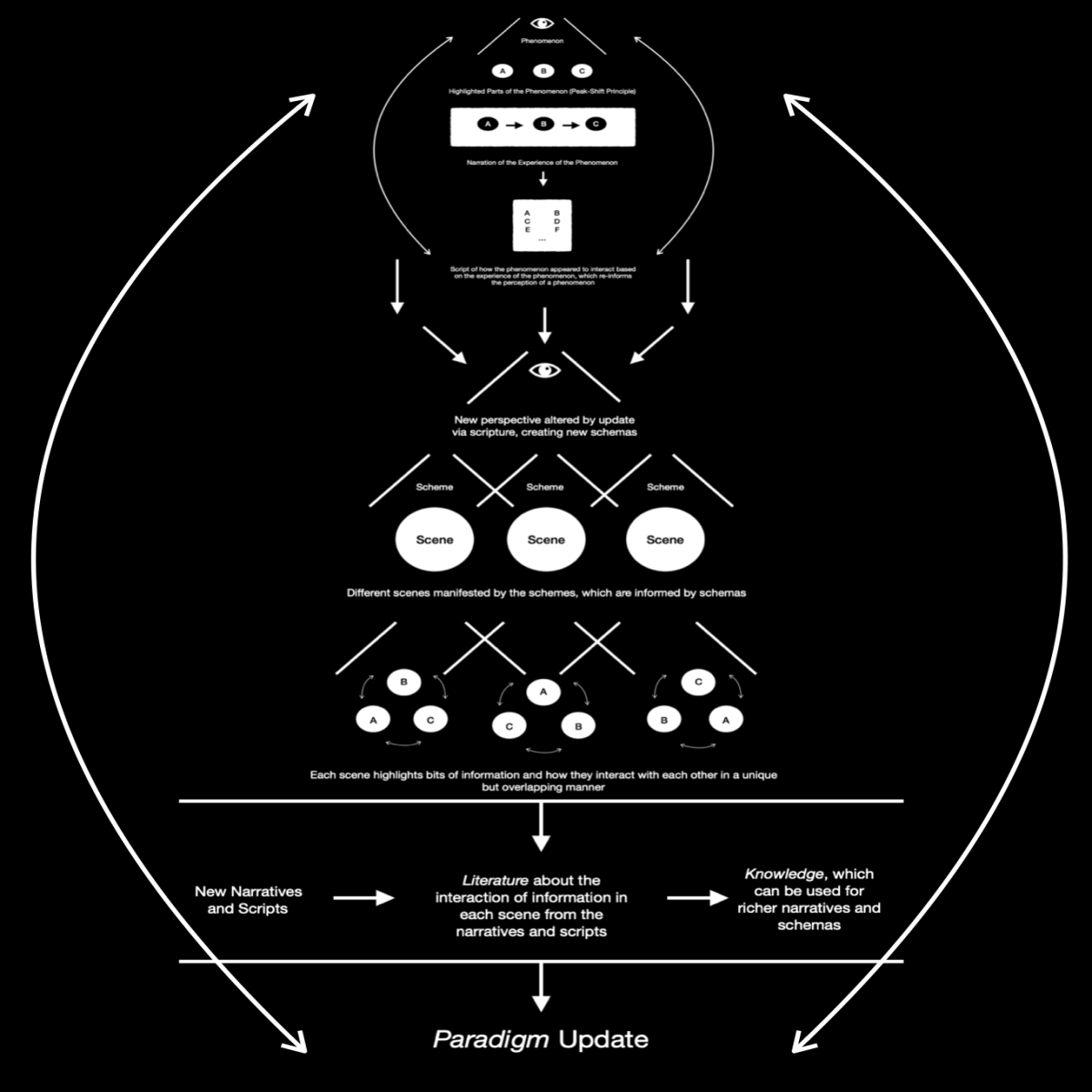
Fig. 4
Figure 4 completes the whole picture that Figure, 1, 2, and 3 begin. In Figure 4, the reader should be able to see the reciprocal nature of the entire model. This effect should be similar to the HSFC model and the SLAM model (10. & 11.). These models work off feedforward and feedback conceptions of cognition. Essentially, this model ensures that the individual is not just working off slow, feedback principles, but can also produce their own models and thus effect change without necessarily having to learn about what they’re doing (feedforward). They can create a kind of Frankenstein’s monster out of past experiences, recruit those different experiences from various areas of the brain (e.g., pre-motor area), and impose previous patterns of behavior from analogous experiences onto the novel experience, which is similar to how metaphor works (12. & 13.).
How is the Paradigmatic Model Useful?
Hell can be defined as a kind of chaos that results from the improper structuring of our behavior. Thus, to avoid hell, we need a kind of structure, a paradigmatic structure to help us move throughout our lives. If the model I am proposing is sound, narrative would be a vehicle for this proper structuring. Importantly, because this model provides an observer and narrator a way to experience a variety of scenes, modes of reality, and thus stimuli in reality, he can compare and contrast viewpoints and positions to make more accurate predictions about the validity of any scheme, schema, or the narrative from which one acquires that schema (14., 15., 16., & 17.). Different schemas may also result in the revitalizing of dead notions via contagion effects, which would help to remodel and restructure the scenes we’ve created for different aspects of reality. By inspiring other works, new viewpoints can be accessed, disseminated, and potentially new and valid knowledge acquired (18.). Through incubation, the identification of problems from narratives may also help to engage problem-solving mechanisms through a process known as incubation, which has been shown to improve problem-solving (19., 20., & 21.). This could help a person avoid what is tantamount to scandals or traps by allowing them to experience those scandals or traps before they encounter them, within a narrative, thinking on them (incubating), and then seeking self-made solutions to those problems if they believe the character’s solution was insufficient via their own experiences.
However, the ability of a person to use this model will most likely be mediated by how creative they are and how intelligent they are (22.). At least, intelligent people may be more likely to effectively make use of word-play, semantics, or metaphor (12. & 23.). This would enable more intelligent people to form connections, useful connections, as long as they were not too separated from the main idea (core) of a situation. If they were too separated from the core of a situation, they would likely have flatter semantic networks (24. & 25.), which would decrease the utility of the novel ideas that they would be able to generate (26.). The generation of more novel and appropriate ideas would be supportive of more efficient paradigmatic updates, which would enable both creative and intelligent people to see reality more clearly than someone who was neither creative nor intelligent. However, intelligence would only help to a certain degree (27.). If a problem requires a sufficient amount of divergent thinking, if the core was unclear, intelligence will no longer be the deciding factor (28. & 29.). I.e., when a problem requires an individual or group of people to produce a solution that is sufficiently unique and appropriate to distinguish itself from a copious amount of solutions that have been shown to be inadequate, the creative individual will most likely have the best solution.
How Might the Brain Enable or Support the Paradigmatic Model?
Because this is not the place for a thoroughly detailed explanation of all of the cytoarchitectonic processes involved in this model, and for the sake of time, I will try to be brief with the areas that may, at least, be involved in the proper functioning of this model.
The area I will begin with will be our visual system. To put it mildly, the visual network is complex. We begin to process stimuli via the retina (30.), which can then be modulated by other stimuli, e.g., auditory stimuli (31.), which may cause physical reactions, depending on the conception of the stimuli (32.). Visual information may be sent to different areas of the brain via the thalamus (33.), which then creates feedback communications (34. & 35), which can lead to different representations of visual or sensory stimuli in three-dimensional space (36.). Our visual system also enables binocular perception, which leads to dual processing of visual information (37.). The visual information we process can also be altered based on the context within which we see it (38.). Lastly, the visual or sensory information we process may also be broken down spatially, as a particular (what) or an object in space where (39.), or even based on color (41.). Regardless, the visual system is exceedingly complex and is still being explored (40.).
This is a seriously truncated description of how we process one kind of sensory stimuli (there are at least four others), which give rise to phenomenon. Nonetheless, it helps to highlight the fact that underlying the fact that we see things is a process of how we see things. If this process is altered for any reason, lesions or insult, or even by our own volition, our conception of sensory stimuli may also be altered. Of course, this is suppositional, but that is why this paper is exploratory rather than confirmatory. More importantly, this brief description highlights the fact that our understanding of sensory phenomenon is contingent upon our ability to perceive the stimulus producing the sensory experience. If there is no stimulus or even the effects of one, there is nothing to be grasped with the senses; you cannot grasp information from any such stimuli. Thus, knowledge is contingent upon our sensory experiences.
Secondly, how we narrate. The cortical organization of speech processing is facilitated by areas such as the temporal lobe, the prefrontal cortex (particularly BA 44), the pre-motor area, the temporal-parietal junction, and the anterior insula (42.). The activation of the language areas are also left-hemisphere dominant (43.). Networks throughout the brain also help to facilitate creativity, such as the FPN (Frontoparietal Network) and DMN (Default Mode Network) (44.). Importantly, verbal creativity was negatively correlated with connectivity in the medial prefrontal cortex of the aDMN (anterior Default Mode Network). The FPN helped to mediate activity in the DMN, and aDMN to facilitate creativity. This allows for functional connectivity to facilitate creativity via multiple regions of the brain to generate narratives, not just areas in the brain involved with speech production. This capacity for greater neural connectivity enables richer narratives, supported by encoded information within different areas of the brain. The main areas of the DMN that enable this process are the PCC (Posterior Cingulate Cortex/Precuneus and the mPFC (medial prefrontal cortex). Another network that may facilitate speech and thus narrative production would be the DMN and ECN (Executive Control Network), whose interactions are mediated early in the creative process by the SN (Salience Network) (46.). The connection between the DMN and ECN, which affects areas such as the IPS (Intraparietal Sulcus), helps to facilitate the production of metaphor. The IFG and the DMN, together with other networks (e.g., the ECN, FPN, and SN), also help to facilitate creative narrative production (47.). However, speech must also be processed. Speech is processed through the comprehension of language (ventral stream) and acoustically (dorsal stream) (45). Thus, narrative production is both region-specific and a broad network process. In this way, when studying how the paradigmatic structure model affects and is affected by the brain, it is important to not focus on too specific an area or to not be sufficiently precise with what kind of work is being produced and studied (e.g., verbally creative works vs., visually creative works); don’t mistake the forest for the trees and vice versa.
To avoid getting caught in the weeds of the neuroscientific literature on the production of speech and narrative, I think this process should be concisely described. In short, we use language to describe our world by accessing a variety of different areas in a networked manner, creating linguistic, paradigmatic structures that then serve as feed-forward models (e.g., HSFC and SLAM) to explore and explain our world for the sake of maintaining a social unit (10., 11., & 22.).
But how do we understand other people’s stories? Mirror neurons. Mirror Neurons can be observed in monkeys and humans alike, who learn by watching others (49.). Areas like BA 44 help to facilitate mirror neuron activity, which is an area that’s also involved in speech production (50.). In fact, we do not only activate BA 44 when producing speech but also when we observe another person reading (51.). Mirror neuron activity also occurs in the ventral Pre-Motor area (PMv), an area of the precentral gyrus near the superior frontal sulcus (SFS), extending into varying parietal areas, when observing others (52.). Activation occurs in these areas and others to facilitate the comprehension of other peoples’ behaviors (e.g., motor behaviors), which are essential to communicating with others (53.).
For example, understanding others, and mimicking their behaviors, helps to facilitate socialization (54.). The frontal lobe (FL), medial temporal lobe (mTL), and the hippocampus also exhibit mirror neuron-like activity, indicating that multiple areas of the brain are involved in comprehending the behaviors of others and to learn from them (55.). The inferior frontal cortex (iFC) plays another role in social cognition (BA 44 and parts of BA 6) (56.), highlighting the importance of mirror neurons in linguistic production and comprehension, and thus a key role for narrative and language in socialization and social learning. But we do not simply learn about others from their movements or what they say, we also learn about others from how they emote. For example, we learn about what causes others pain from how they respond to pain (57. & 58.), or when someone is around something or smells something that is repulsive to them (59.), when others make mistakes (60.), and when someone else is given a reward, which activates the nucleus accumbens and the ventral striatum (61.). Interestingly, our ability to make predictions about the judgments of others is also facilitated by how similar they are to us, which may be the root of the perennially potent phrase, “Know thy self” (62.).
Together, these findings clearly suggest that we learn by observing others, what they do, and how they respond. The overlap between the mirror neuron networks and the linguistic networks suggest that narratives, scripts, and literature emerge from underlying neural mechanisms that have helped to facilitate learning by humans throughout our phylogenetic, evolutionary history, and thus would help us learn today. This adds credence to the notion that the model of paradigmatic structures helps to facilitate learning and may improve the lives of people who make use of it by taking advantage of the power that narratives provide us.
But such information can not only facilitate individual learning, but also social learning, sometimes recognized as transpersonal learning. It is likely that this is produced by the mechanisms that enable us to engage in Theory-of-Mind (ToM) thinking.
Theory-of-Mind (ToM) is the ability to understand that the mental states of others are different from our own. ToM also facilitates our understanding of the feelings, goals, and intentions of others. Humans are not born with ToM and it appears that most of us develop ToM late in our pre-school years (63.). A Meta-Analysis of ToM indicates that the areas involved in the process include the temporoparietal junction (TPJ) the superior temporal lobe (ST) and the frontal lobe (FL) (64.). The keen reader should notice that some of these areas or portions of these areas overlap with linguistic areas and narrative production. These areas are partially involved when recognizing the intentionally of others (65.). For example, a Mind-in-the-Eyes task helps us to recognize the emotional or sentimental mental states of others, the IFC and TPJ (66.). False belief tasks, which enable an observer to discriminate between true and false statements of belief through the behaviors of others, activates the IFC, TPJ, and Precuneus (67.). Some of these areas are also activated during narrative production, like the precuneus, which is involved in divergent thinking and idea generation. These studies highlight that the TPJ serves as a nexus for understanding the mental states of others, their feelings, goals, and intentions (68.). Of note, judgments about social animacy, which relate to our ability to learn about others (recall mirror neurons), primarily involve the inferior frontal cortex (IFC), or in other words, BA 44, which again, is related to narrative production.
Empathy is also essential for understanding the mental states of others, beyond just understanding that others have intents and goals that are mediated by their sentiments, or the ability to project our personality into others. For example, in a study on empathetic accuracy, Zaki et al., 2009 studied how well a viewer’s estimation of a performer’s emotional state matched what the performer stated their emotional state was. This task activated the iPL (inferior parietal lobe) and the mPFC (medial prefrontal cortex) (70.). Medial prefrontal cortex activity also increased when observers viewed the social ostracization of strangers compared to their friends (71.). In another study, those who engaged in more helpful behavior showed greater mPFC activity when shown images of strangers in need (72.). This activity likely occurs because it enables us to predict the mental states of others and thus to track their behavior over time; it is also thought that this behavior helps us to engage in more prosocial behavior by understanding the mental states of others (73. & 74.). Interestingly, activity in the dorsomedial prefrontal cortex (dmPFC) also is observed in metaphor production (69. & 48.). In relation to what is presented here about mirror neurons and why we empathize, one might theorize that metaphors enable us to elicit the mental states of others, from the minds of others, via their reaction to the metaphor, to help us understand them.
When we think about others, we also have to differentiate ourselves from them. Denny et al., 2012 showed that areas of the brain involved in social judgment and personal judgment overlap, and include the mPFC and TPJ (75.). Of note, judgments about the self are more ventral while judgments about others are more dorsal (vmPFC and dmPFC respectively). One way to think about this is as if we are producing judgments about the self (vmPFC) and producing judgments to comprehend others (dmPFC) (45.). Individuals also seem to attribute their actions to social constraints while attributing the actions of others to their inherent personalities. This interesting facet of self vs. social personal judgments is known as fundamental attribution error (76.). Judgments about ourselves are also temporally dependent. For example, judgments about our future or past selves produced mPFC action that was similar to the type of activation that occurs when we think about others (77. & 78.). I.e., when we make judgments about where we were in the past compared to where we currently are and where we can be in the future, we are seeing ourselves as a different person or character (79.).
What this helps to show is that areas involved in ToM, Empathy, and Personal and Transpersonal Judgments involve areas of the brain that overlap with narrative production. This is highly suggestive of the fact that, when taken into consideration with mirror neuron activation, stories enable us to learn about ourselves and others through written or spoken narrative. By learning about others, we avoid their mistakes, can become more prosocial, and more willing to help others. In theory, literary mechanisms such as metaphor also enable us to elicit information from our social environment by making novel connections about stimuli within our environment, which enables us to learn more about others and the environment.
To understand how we are alerted to relevant stimuli, which provide us with phenomenological experiences, why we are motivated to engage in certain behaviors and not others, how we integrate our feelings into our actions and choices, all of which are extremely relevant to understanding the human condition and narrative, it is important to understand several aspects of our limbic system.
What is it in our brains that arouses us? To put it simply, the Amygdala. The amygdala is relevant for the detection of emotional information and rapid responses to that information. Thus the amygdala is relevant for learning the emotional significance of that information (80.). The amygdala is broken down into three parts. The Basolateral nuclei of the amygdala connect to the hippocampus and pFC, which thus play a relevant role in learning, while the central nucleus and cotricomedial nuclei are connected to the hypothalamus, and other regions involved in autonomic response, which enable the amygdala to alter our response to those autonomic signals (81.). When the amygdala is damaged, unusual behavior is usually followed, sometimes called psychic blindness (82. & 83.). For example, lesions of the amygdala cause individuals to have trouble identifying fearful expressions and faces (84. & 85.). Individuals with amygdala lesions also are extremely trustworthy (86.), i.e., they have impaired threat detection (87.). Our amygdalae also respond to phobia-inducing stimuli (88. & 89.), which can be triggered even when the stimulus is outside conscious awareness (90.). Together, these findings suggest that the amygdala is involved in emotional learning via, at least, fear conditioning (91.). In one study, damage to the amygdala prevented autonomic responses, while hippocampal damage prevented the memorization, or learning of what caused the autonomic response (92.). This study confirms the idea that the amygdala is responsible for alerting us to relevant stimuli within our environment.
Importantly, we do not have to physically learn to avoid or to be alert around a stimulus or set of stimuli. Rather, we can learn to avoid or be alert to certain stimuli through language and symbol (93. & 94.). The amygdala also enables an individual to learn via reward-based learning, i.e., an individual becomes alerted by the amygdala when they are in the presence of something that is rewarding, drawing their attention towards that stimulus (95.). Another study showed that the amygdala is more responsive to stimuli intensity than valence (whether it is scary or alluring) (96. & 97.), which means that the amygdala is involved with emotional learning in so far as it helps us to focus our attention on relevant stimuli (98. & 99.). This results in emotional encoding, which occurs via interactions between the hippocampus and the amygdala (100.). This is confirmed by the fact that the amygdala plays a role in the memorization of events that are emotionally charged (101. & 102.). I.e., the more emotionally intense an experience, the easier it is to recall and remember it. This finding is supported by the fact that the amygdala reduces memory extinction; intense emotional memories are harder to forget because they are coded by the hippocampus as being highly relevant memories (103.).
For the sake of my model, the amygdala should enable anyone exploring narrative, literature, scripture, or scripts to recognize when a character, set of characters, or the receiver of the narrative (etc.) is alerted to something, or that they should or might want to be alerted to something because there is probably something worth learning about. This goes for writers of scripts and narratives as well, or for anyone exploring any type of literature. Amygdala activity will alert us to relevant information, perhaps even based on our past experiences, given the association between the amygdala and the hippocampus. Thus, the amygdala modulates what we pick up on in narrative; alerts us to relevant information within narratives; and at least, facilitates our understanding of how it causes us to become alerted to information, allowing us to theorize and predict the mental states and behaviors of individuals and characters within any given narrative. This last point is especially true if they are obviously alerted to something relevant within their environment, or are acting oddly given their circumstances.
But how do we know if a stimulus is rewarding or not? The ventral striatum, nucleus accumbes (NA), and the dopaminergic pathway are the areas involved in reward and motivation, and thus help us recognize when a stimulus is rewarding or not. Specifically, the NA is involved in wanting a stimulus (104.), while the NA-shell is involved in the feeling of pleasure when we acquire a stimulus or achieve a goal (105.). The NA specifically activates when one unexpectedly receives a reward via a stimulus (107.), and is also involved in making predictions about a stimulus or goal (106.). This means that the NA responds to rewards and then tracks the pattern that produced the reward; it wants and then senses whether a stimulus was rewarding or not based on past experiences with that stimulus (108.). I.e., the NA conditions you to seek out your favorite treat, chocolate, coffee, tobacco, alcohol, marijuana, or at least, cocaine; it lays the groundwork for achieving and acquiring goals.
In relation to my model, our understanding of the reward and pleasure centers of cognition, which also involve areas like the ventral tegmental area, which tracks the activity of the NA via dopaminergic activity in the striatum, helps us to understand why people seek out certain rewards or engage in goal-oriented behavior – they get something out of it: pleasure. It is thus relevant for individuals to incorporate pleasure, what drives pleasure, how it can be hijacked with imagery, and how pleasure and reward shape our behavior, all into their understanding of people, narratives, scripts, scripture, and literature to get a fuller understanding of people within those narratives (etc.). Once again, by understanding reward-seeking or pleasure-seeking behaviors, we can make more accurate predictions about the behaviors of others, or understand the mental states of others, both of which increase one’s ability to interact with them, help them, or to identify the tracts in their lives that lead them astray, which one would be wise to avoid.
Implied in my description of the NA and the Amygdala is the notion that our emotions are transformed into action. The area involved in this transformation of sentiment or mere emotion into full-fledged action is the cingulate cortex. The anterior cingulate cortex (ACC), for example, is involved in selecting motor actions, considering the cost and effort (NA activity) entailed in taking those actions, and weighing how much reward is to be gained by following through with a set of actions via information gained from experiences from similar actions in the past (109.). The ACC also integrates affective signals with cognitive control to increase one’s confidence in their actions when they are unsure about what they’re going to do (110.). The cingulate also tracks pain (111.). More precisely, some areas of the cingulate track the presence or absence of pain, while others respond to the intensity of the painful stimulus (112.). The anterior cingulate is also responsible for emotional processing (113. & 114.). The cingulate is connected to the amygdala (how stimulating is the stimulus), hypothalamus (is there pain, e.g.,), insula (how does it feel., taste, pain, is something pleasant or unpleasant), and orbitofrontal cortex (is one’s behavior appropriate), which means that this area is activated by and incorporates sensory and emotional information from these areas into its determinations about whether an action is worth taking or not. This is especially true for signals received from the autonomic nervous system (115. & 116.). Interestingly, during cognitive tasks, activity in the rostral portion of the cingulate decreases while increasing in the dorsal portion; and for emotional tasks, activity in the rostral portion of the cingulate increases while decreasing in the dorsal portion of the cingulate (117. & 118.). In one study, the midportion of the cingulate, the midcingulate, was activated by negative emotions, pain, and cognitive control, highlighting the role that the cingulate plays in integrating such information (110.). In summation, this means that the cingulate cortex is a mediator between emotional signals and cognitive control (119.); the cingulate links emotion and cognition and affects the behaviors which emerge from both.
For the model, an understanding of cingulate activity highlights the relevancy of emotion in the behavior of characters or individuals within any narrative. An understanding of cingulate activity may give us an understanding of how a character goes forward with or decides to avoid certain behaviors or actions; like whether one should ride a skateboard given that he’s fallen down from one every time he’s tried to ride one. More interestingly, such an example may cause one to ask why one would continue to try to ride a skateboard given that he has fallen down from one every time he’s tried to ride one. In other words, it gives structure to the absurdity of human activity. When trying to understand a narrative, Scripture, a script, or literature, understanding the experiences of the people in those stories, how those experiences shaped their behavior, and then making predictions about their behaviors in the future based on who they are and what they’ve been shaped by, enables us to understand their mental states, which may help us to understand ourselves. This theoretical process enables us to more efficiently learn about a character or set of characters in a narrative by incorporating the seemingly absurd into their behavioral processes; it also enables us to avoid their mistakes by feeling their pain (recall the discussion on Empathy and Mirror Neuron activity) or more richly conceptualizing their pain.
The final portion of my discussion on the areas of the limbic system that I think are relevant to the paradigmatic model I’m proposing is the Orbitofrontal Cortex (OFC) or ventral medial prefrontal cortex (vmPFC). As noted above, emotional signals are critical for effectively guiding ourselves toward outcomes that benefit us and away from outcomes that will cause us harm (120.). The OFC enables this by attributing value to rewards and punishments while using an understanding of rewards and punishments to guide adaptive behaviors (which is mediated by the cingulate) (121.). Individuals with OFC damage engage in inappropriate behavior, which suggests that the OFC or vmPFC is the neural substrate responsible for moral behavior (122. & 123.). This is indicated by the fact that individuals with OFC or vmPFC damage often fail to empathize with others, which has been observed in violent criminals (124.). People with OFC damage also perform poorly on tasks that require an individual to weigh past losses and gains to make appropriate choices about the future and present (125. & 126.). It is suggested that, if not innate, these issues may arise from substance dependence, as substance dependence is linked with atypical activity in the OFC (127. & 128.). For example, healthy OFC activity will indicate to a person when food becomes less desirable or satiating (129.). Because substance dependence is related to an “infinite thirst” for a particular substance, this feeling of satiation for the substance does not exist or is grievously impaired in the substance-dependent individual. Together, these findings indicate that the OFC is especially important for learning in situations that require an individual to change according to corresponding patterns of reward or punishment. This means that the OFC or vmPFC allows us to represent the costs and benefits associated with any choice, which enables more effective decision-making (130.). Interestingly, patients with OFC damage do not feel regret, i.e., they feel no sense of responsibility for their actions (131.). This suggests that the OFC is responsible for the abstract notion of accountability, or responsibility. This is supported by a study which showed that OFC activity is tied to situations in which participants feel agency for their choices, rather than when an outcome occurred as a matter of circumstance or chance (132.).
Because the goal of my model is to support healthy behaviors and decision-making, the OFC plays a crucial role in facilitating this process. When we are presented with a narrative, which enables us to extract a lesson from it, and we are given two scenarios, one of which will be punishing and the other of which will be rewarding, it is the OFC that will help to facilitate which one we choose. Once again, an understanding of this aspect of our decision making and limbic systems allows us to make better predictions about our own behaviors, the behaviors of others within narratives, the interactions of characters within narratives, all of which enable us to engage in prosocial behavior for the understanding we acquire from the mental states and emotional states of others. This is once again facilitated by our empathic network and mirror neurons. The knowledge acquired via prediction and validation of the behaviors and choices of characters within any narrative facilitates learning, which enables us to alter our own scripts and narratives to avoid the pitfalls of others or to take advantage of the choices that they did, especially if their choices are consistently rewarding and sustaining over a very long period of time; the latter of which is precisely what the addict or substance-dependent individual is seeking, only in an erroneous manner.
But how exactly do we learn, what facilitates our ability to learn? At least, the hippocampus and the basal ganglia (BG), which I have referenced previously by name (the hippocampus) and in another form in my discussion of reward-seeking behavior (the striatum). To start, the BG. Much has been gleaned from patients with Parkinson’s and Huntington’s disease about the BG. For example, patients with impaired BGs cannot keep a reticule on a target as well as those with unimpaired or undamaged BGs (133.). Such individuals also have great trouble with learning sequencing (134.). This is supported by the fact that improvement in sequence learning is facilitated by the striatum (135.). Individuals with impaired or damaged BGs also have trouble learning written scripts that are reflected in a mirror (136.) and have trouble with probabilistic learning (137.). Taken together, these findings suggest that the BG is involved in implicit and procedural learning (138.). For example, one study identified that the BG is involved in error-driven learning via the dopaminergic system (139.); i.e., individuals learn from action-outcome associations, otherwise known as implicit learning.
But how do these memories get stored and accessed, i.e., learned? The Hippocampus. The hippocampus plays a role in memory consolidation, i.e., the process by which memories are strengthened for long-term retention (140.). This can potentially happen in two different ways. The hippocampus can encode memories via the strengthening of the bonds of any two memories over a long period of time (141.). This suggests that the hippocampus helps to encode these memories over the long period of time (142.), rather than immediately or in a relatively shorter period of time. However, multiple trace theory suggests that memory consolidation need not occur over great periods of time, but can happen rapidly (143.). I.e., every time a memory is retrieved, the hippocampus strengthens that memory, or a new trace is indexed to that event. Regardless of whether the process of memorization occurs rapidly or incrementally over a long period of time, which may seem inefficient, the hippocampus is evidently responsible for encoding information into memory. Importantly, anterior portions of the hippocampus are involved in encoding and posterior in retrieval (144. & 145). The hippocampus is involved in retrieval by linking one component of a memory together with another one (146.). This process is known as pattern completion, wherein one smaller piece of information can be used to constitute the whole (147.). Recollection or retrieval of one of these smaller pieces of information relies on something like familiarity (148. & 149.). Retrieval by familiarity relies on areas near the hippocampus like the perirhinal cortex and the dorsal medial nucleus (150). Importantly, this process of retrieval also enables the hippocampus to be involved in processes like imagining the future or future planning (151.). I.e., the ability to remember the past improves the ability to plan for the future (152). The hippocampus enables this by binding pieces of information together in novel ways to aid with problem-solving and to prepare for future actions and scenarios (153.).
Together, the BG and the hippocampus help to facilitate predictive learning by procedure and implication and to encode and retrieve information for problem-solving and future scenarios and activities. In relation to my model, healthy functioning of the BG or the hippocampus enables an individual to extract the implicit lesson from a narrative, encode that information into memory, piece together different lessons from one or more scenarios, and engage in problem-solving and future planning for themselves or for characters within their own narratives. All of this is abetted by a rich understanding of the emotions, desires, and feelings of others and how they affect their behavior; emotional comprehension is facilitated by a thorough ToM, understanding of empathy, and mirror neuron network; these networks also have significant overlap with our linguistic production and comprehension networks; lastly, all of these are manifested and informed by the phenomenon of our senses. In other words, my model is coherent with, and helps to inform, the neuroscientific literature.
Discussion
I think I have sufficiently laid out the linguistic framework, its structural model, the neuroscientific literature supporting the model, and their relationship to the paradigmatic structure of narrative. But to lay out the argument once more: via our senses, we have phenomenological experiences made of distinct parts, which when put into their proper context, form narratives, and when those narratives are put into a script, form written narratives, or scripture. From scripture, we get schemas, which provide us with a varied set of schemes, that produce different scenes. Each scene enables an observer or a collection of observes to simultaneously experience a collection of phenomenological experiences in a distinct and overlapping manner. The scripts and narratives that are informed by the interaction of these scenes can be called literature, as they provide knowledge about the phenomenon constrained by the schema, via any given scheme or scene that is provided by the schema or schemas. A schema, and the literature that results from them, may claim to have access to knowledge if predictions that emerge from the scriptural literature are predictive of phenomenological experiences within reality. This will either provide an individual or group of people success or failure, which is the ultimate judge of the schema that emerges from the narrative, not any internal set of rules, which are subject to fallacious, circular thinking. All of this is supported by the neurological literature, and thus is a product of our biology.
Because it is a product of our biology, this creates constraints upon which models are likely to be deemed valid and invalid. Models that acknowledge and build upon a continually evolving understanding of biological knowledge will be fitter, in the BVSR sense (154). As is implied in some of the literature in this paper, and because our biological structures are a product of this process, fitter narrative models will also have to acknowledge our evolutionary history, and will thus be judged by whether they acknowledge and work from our evolutionary history or ignore it. The models that incorporate knowledge about Humanity’s evolutionary history will likely be fitter than those that do not. Lastly, because value can be thought of as that which is conducive to the long-term success, or fitness (in the Darwinian sense), of people within and without as many environments as possible (155.), narratives that have lasted the longest and provided the most stable structures for a group of people should also be a constraining factor; i.e., foundational narratives are sounder than newer narratives, as they simply have greater value; there’s something to them if they’ve lasted that long. Together, foundational narratives, scripture, biological frameworks, and evolutionary theories -- all of which are highly predictive and thus provide valid knowledge of phenomenological experiences of the world and its constituent parts -- are limiting and constraining factors for which narratives should be explored and which one’s should not.
In other words, you don’t get to write, create, or do whatever you want. You are and ought to be limited by your biological form, your evolutionary history, and the foundational narratives you’ve inherited, all of which have gotten you to where you are today, which without, you would not be here. From a creative standpoint, this is fantastic, as constraint is highly conducive to creative achievement, more so than free-reign or unbounded limitation, which is more likely to produce less creative work (156. & 157.). However, this also means that any given group of people cannot simply tolerate whatever comes across their path, especially if it proposes narratives and schemas that are antithetical to the Scriptures of any given people, the foundational beliefs of those people; knowledge acquired through the study of biology, which has proven to be valid; or at least, sound knowledge acquired through predictive models derived from evolutionary theories.
Because these structures provided the natural and anthropogenically derived spaces wherein we reside, for which not one of us is the cause, it is exceedingly unwise to undermine or ignore any of these structures. A society or individual does so at the peril of being thrown into a kind of literal hell, if not the actual one for doing so. To try to dismantle any of these structures will likely result in them reasserting themselves in truly bizarre ways, which is an aspect of our and our world’s nature, which is to achieve a kind of homeostasis or equilibrium (158.). In this way, a people who abandon or ignore the sanctity of these structures will likely be judged by them, ultimately judged if you will, for who knows if they’d survive the upheaval wrought by their foolhardiness. This is partially borne out by the fact that the predictions we make from the literature we derive from the schemas created by our narratives provide us with either valid or invalid conclusions. If you’ve accepted this premise, the premise of the ultimate judgment, which results from abandoning at least the evolutionary structure, biological structure, and foundational structure provided to an individual by their society’s holy script, must follow; you can’t wriggle out of it.
Conclusion
In this paper, I have explored the paradigmatic model, the terminology that gives the model structure, its relationship to our neural architecture, and how the model can help us improve our lives by enabling problem-solving through narrative. Because this model is based on a biological substrate, is confirmed by that substrate, and because that biological substrate emerges from our evolutionary history, it is necessarily bound to that biological substrate and evolutionary history, and thus so too is narrative. Because Humans also reside within a structure that enables them to engage in complex socialization, for which none of them are the cause, to continue to engage in that complex socialization, they are also bound to those foundational structures and to the foundational beliefs of those anthropogenic structures. Because some of those structures are antithetical to each other, this necessitates a kind of internal intolerance from the structure to maintain the complex functioning of that structure. Our understanding of these structures, and our ability to properly transform these structures for the betterment of Mankind is informed by the paradigmatic model I’m proposing; which again, necessitates working within, at least, the three spaces listed in the discussion section of this paper.
In conclusion, this exploratory model should be informative for and informed by neuroscientific research into narrative; literary studies on contemporary and classical works; and may help an individual structure their lives to avoid living in a disorganized and costly fashion.
Bibliography
- The Science of Art – A Neurological Theory of Aesthetic Experience, Ramachandran and Herstein, 1999
- Representational change and analogy: How analogical inferences alter target representations, Blanchette and Dunbar, 2002
- Expert music performance: Cognitive, neural, and developmental bases, Brown, Zatoree, Penhune, 2015
- Neural correlates of verbal creativity: differences in resting-state functional connectivity associated with expertise in creative writing, Lotze et al., 2014
- Professional training in creative writing is associated with enhanced fronto-striatal activity in a literary text continuation task, Erhard et al., 2014
- Expertise in Musical Improvisation and Creativity: The Mediation of Idea Evaluation, Klienmintz et al., 2014
- Neuro-intelligence, neuro-metrics and the next phase of brain imaging studies, Haier, 2009
- A Report on the 40-Year Follow-Up of the Torrance Tests of Creative Thinking: Alive and Well in the New Millennium, Cramond et al., 2005
- The Creativity Maze: Exploring Creativity in Screenplay Writing, Bourgeois-Bougrine, Glaveanu, Botella, 2014
- Computational neuroanatomy of speech production, Hickok, 2017
- Bridging computational approaches to speech production: The semantic–lexical–auditory–motor model (SLAM), Hickok, 2015
- Verbal creativity in autism: comprehension and generation of metaphoric language in high-functioning autism spectrum disorder and typical development, Kasirer and Marshal, 2014
- Creating metaphors: The neural basis of figurative language production, Benedek, 2014
- The proactive brain: using analogies and associations to generate predictions, Bar, 2007
- Human creativity, evolutionary algorithms, and predictive representations: The mechanics of thought trials, Dietrich & Haider, 2015
- Prediction, cognition and the brain, Bubic, Cramon, Schuboltz, 2010
- The proactive brain: using analogies and associations to generate predictions, Bar, 2007
- Writer–Reader Contagion of Inspiration and Related States: Conditional Process Analyses Within a Cross-Classified Writer × Reader Framework, Thrash et al., 2016
- Does Incubation Enhance Problem Solving? A Meta-Analytic Review, Sio and Ormerod, 2009
- Incubation and intuition in creative problem solving, Gilhooly, 2016
- Problem-solving and creativity: insights from students’ individual composing pathways, Burnard, 2004
- Evolution, creativity, intelligence, and madness: “Here Be Dragons”, Jung, 2014
- Making creative metaphors: The importance of fluid intelligence for creative thought, Silvia and Beaty, 2012
- Investigating the structure of semantic networks in low and high creative persons, Kenett, Anki, and Faust, 2014
- Loose but Normal: A Semantic Association Study, Mohr et al., 2005
- The Standard Definition of Creativity, Runco and Jaeger, 2012
- Relationship of intelligence and creativity in gifted and non-gifted students: An investigation of threshold theory, Preckel, Holling, and Weise, 2006
- The relationship between intelligence and creativity: New support for the threshold hypothesis by means of empirical breakpoint detection – Juak, Benedek, Dunst, and Neubauer, 2013
- Is creativity without intelligence possible? A Necessary Condition Analysis – Krakowski, et al., 2016
- The new visual neurosciences, Werner and Chalupa, 2014
- Multisensory response modulation in the superficial layers of the superior colliculus, Ghose, et al., 2014
- Signals conveyed in the pulvinar pathway from superior colliculus to cortical area MT, Berman and Wurtz, 2011
- Superior colliculus and visual attention, Krauzlis, 2014
- Guarding the Gateway to Cortex: Attention in Visual Thalamus, McAlonan et al., 2008
- Attention modulates responses in the human lateral geniculate nucleus, O’Connor et al., 2002
- Whiter the Hypercolumn, Ts’o, 2009
- The cortical organization of binocular vision, Freeman, 2014
- Attention determines contextual enhancement versus suppression in human primary visual cortex, Flevaris and Murray, 2015
- Ventral and dorsal cortical processing streams, Bell et al., 2014
- Felleman, Distributed hierarchical processing in the primate cerebral cortex, 1991
- The processing of color in extrastriate cortex, Gegenfurtner and Kiper, 2004
- The cortical organization of speech processing, Hickock and Poeppel, 2007
- The Brain Basis of Language Processing: From Structure to Function, Friederici, 2011
- Common and Distinct Brain Networks Underlying Verbal and Visual Creativity, Zhu et al., 2017
- Neural Basis of Speech Perception, Hickok and Poeppel, 2016
- The Association between Resting Functional Connectivity and Creativity Takeuchi et al., 2012
- Creativity and the default network: A functional connectivity analysis of the creative brain at rest, Beaty et al., 2014
- Brain networks underlying novel metaphor production, Beaty, Silvia, Benedek, 2017
- The mirror mechanism: recent findings and perspectives, Rizzolatti and Fogassi, 2014
- Cortical mechanisms of human imitation, Iacoboni et al., 1999
- Neural circuits involved in the recognition of actions performed by nonconspecifics: an fMRI Study, Buccino et al., 2004
- Brain regions with mirror properties: A meta-analysis of 125 human fMRI studies, Molenberghs et al., 2012
- Language within our grasp, Rizzolatti and Arbib, 1998
- The antecedents and consequences of human behavioral mimicry, Chartrand and Lakin, 2013
- Single-neuron responses in humans during execution and observation of actions, Mukamel et al., 2010
- Evidence of mirror neurons in human inferior frontal gyrus, Kilner, 2009
- How do we perceive the pain of others? Window into the neural processes involved in empathy, Jackson et al., 2005
- Empathy for pain involves the affective but not sensory components of pain, Singer et al., 2004
- Both of us disgusted in my insula: the common neural basis of seeing and feeling disgust, Wicker et al., 2003
- Modulation of activity in medial frontal and motor cortices during error observation, van Schie et al., 2004
- A key role for similarity in vicarious reward, Mobbs et al., 2009
- Perceived similarity and neural mirroring: evidence from vicarious error processing, Carp et al., 2009
- Theory-of-Mind development: Retrospect and prospect, Flavell, 2004
- Fractionating theory of mind: A meta-analysis of functional brain imaging studies, Schurz et al., 2014
- Movement and mind: a functional imaging study of perception and interpretation of complex intentional movement patterns, Castelli et al., 2000
- Social Intelligence in the normal and autistic brain: An fMRI study, Baron-Cohen et al., 1999
- People thinking about thinking people: the role of the temporo-parietal junction in “theory of mind,” Saxe and Kanwisher, 2003
- A nexus model of the temporal parietal junction, Carter and Huettle, 2013
- The Dorsal Medial Prefrontal Cortex Is Recruited by High Construal of Non-social Stimuli, Baetens, Ma, and Overwalle, 2017
- The neural basis of empathic accuracy, Zaki et al., 2009
- Empathy for the social suffering of friends and strangers recruits distinct patterns of brain activation, Meyer et al., 2013
- The neural correlates of empathy: Experience, automaticity, and prosocial behavior, Rameson et al., 2012
- Towards a neuroscience of empathy: ontogeny, phylogeny, brain mechanisms, context and psychopathology, Gonzalez-Liencres et al., 2013
- Toward a cross-species understanding of empathy, Panksepp and Panksepp, 2013
- A meta-analysis of functional neuroimaging studies of self-and other judgments reveals a spatial gradient for mentalizing in medial prefrontal cortex, Denny et al., 2012
- Spontaneous mentalizng predicts the fundamental attribution error, Moran et al., 2014
- Self-reflection across time: Cortical midline structures differentiate between present and past selves, D’Argembeau et al., 2008
- Saving for the future self: neural measures of future self-continuity predict temporal discounting, Ersner-Hershfield et al., 2009
- Modulation of medial prefrontal and inferior parietal cortices when thinking about past, present, and future selves, D’Argembeau et al., 2010
- Emotion and cognition and the amygdala: from “what is it?” to “what’s to be done?”, Pessoa, 2010
- Neuroanatomy of the primate amygdala, Freese and Amaral, 2009
- Amygdala lesions in rhesus macaques decrease attention to threat, Dal Monte et al., 2015
- Impact of amygdala, orbital frontal, or hippocampal lesions on threat avoidance and emotional reactivity in nonhuman primates, Machado et al., 2009
- The enigma of the amygdala: on its contribution to human emotion, Aggleton and Young, 2000
- Amygdala damage impairs emotion recognition from music, Gosselin et al., 2007
- The human amygdala in social judgment, Adolphs et al., 1998
- Impaired threat prioritisation after selective bilateral amygdala lesions, Bach et al., 2015
- Fear is fast in phobic individuals: Amygdala activation in response to fear-relevant stimuli, Larson et al., 2006
- Association between amygdala hyperactivity to harsh faces and severity of social anxiety in generalized social phobia, Phan et al., 2006
- Human amygdala responsivity to masked fearful eye whites, Whalen et al., 2004
- Coming to terms with fear, LeDoux, 2014
- Double dissociation of conditioning and declarative knowledge relative to the amygdala and hippocampus in humans, Bechara et al., 1995
- Activation of the left amygdala to a cognitive representation of fear, Phelps et al., 2001
- Social learning of fear, Olson and Phelps, 2007
- The role of the amygdala in emotional processing: A quantitative meta-analysis of functional neuroimaging studies, Sergerie et al., 2008
- Dissociated neural representations of intensity and valence in human olfaction, Anderson et al., 2003
- Dissociation of neural representation of intensity and affective valuation in human gustation, Small et al., 2003
- The interface between emotion and attention: A review of evidence from psychology and neuroscience, Compton 2003
- How brain beware: Neural mechanisms of emotional attention, Vuilleumier, 2005
- The contextual brain: implications for fear conditioning, extinction and psychopathology, Maren et al., 2013
- Physiological basis for emotional modulation of memory circuits by the amygdala, Paz and Pare, 2013
- The slow forgetting of emotional episodic memories: An emotional binding account, Yonelinas and Ritchey, 2015
- Amygdala-prefrontal synchronization underlies resistance to extinction of aversive memories, Livneh and Paz, 2012
- Parsing Reward, Berridge and Robinson, 2003
- Advances in the neurobiological bases for food “liking” versus “wanting,” Castro and Berridge, 2014
- Anticipation of increasing monetary reward selectively recruits nucleus accumbens, Knutson et al., 2001
- Predictability modulates human brain response to reward, Berns et al., 2001
- Discreate coding of reward probability and uncertainty by dopamine neurons, Fiorillo et al., 2003
- Functional organization of the medial frontal cortex, Rushworth et al., 2007
- The integration of negative affect, pain, and cognitive control in the cingulate cortex, Shackman et al., 2011
- Nociocepetive processing in the human brain, Tracey, 2005
- Dissociable neural responses related to pain intensity, stimulus intensity, and stimulus awareness within the anterior cingulate cortex: A parametric single-trial laser functional magnetic resonance imaging study, Buchel et al., 2002
- Cognitive and emotional influences in anterior cingulate cortex, Bush et al., 2000
- Differential engagement of anterior cingulate cortex subdivisions for cognitive and emotional function, Mohanty et al., 2007
- Anterior cingulate activity during error and autonomic responses, Critchley et al., 2005
- Functional subdivisions within anterior cingulate cortex and their relationship to autonomic nervous system function, Matthews et al., 2004
- Reciprocal suppression of regional cerebral blood flow during emotional versus higher cognitive processes: Implications for interaction between emotion and cognition, Drevets and Raichle, 1998
- Cognition-emotion interactions, Pessoa and Pereira, 2013
- Running on empty: neural signals for self-control failure, Inzlicht and Gustell, 2007
- Descartes’ error: Emotion, reason, and the human brain, Damasio, 1994
- Neurobiology of economic choice: A good-based model, Padoa-Schioppa, 2011
- Insensitivity to future consequences following damage to human prefrontal cortex, Bechara et al., 1994
- Emotion related learning in patients with social and emotional changes associated with frontal lobe damage, Rolls et al., 1994
- The psychopathic magnetized: insights from brain imaging, Anderson and Kiehl, 2012
- Emotion, decision making and the orbitofrontal cortex, Bechara et al., 2000
- Abstract reward and punishment representations in the human orbitofrontal cortex, O’Doherty et al., 2001
- Reduced neural tracking of prediction error in substance-dependent individuals, Tanabe et al., 2013
- Primate orbitofrontal cortex and adaptive behavior, Roberts, 2006
- Activation of the human orbitofrontal cortex to a liquid food stimulus is correlated with its subjective pleasantness, Kringelbach et al., 2003
- The human orbitofrontal cortex: linking reward to hedonic experience, Kringlebach, 2005
- The involvement of the orbitofrontal cortex in the experience of regret, Camille et al., 2004
- Regret and its avoidance: A neuroimaging study of choice behavior, Coricelli et al., 2005
- The impaired learning of semantic knowledge following bilateral medial temporal-lobe resection, Gabrieli, 1998
- Sequential Movement skill in Parkinson’s disease: A state-of-the-art, Ruitenberg et al., 2015
- Learning of sequential finger movement sin man: A combined kinematic and positron emission tomography (PET) study, Seitz and Roland, 1992
- Dissociations between skill learning and verbal recognition in amnesia and dementia, Martone et al., 1984
- Cortico-striatal contributions to feedback-based learning: Converging data from neuroimaging and neuropsychology, Shohamy et al., 2004
- Learning and memory functions of the basal ganglia, Packard and Knowlton, 2002
- A neural substrate of prediction and reward, Schultz et al., 1997
- The consolidation and transformation of memory, Dudai et al., 2015
- The medial temporal lobe, Squire et al., 2004
- Memory consolidation, Squire et al., 2015
- The hippocampal complex contributions to retention and retrieval of recent and remote episodic and semantic memories: Evidence from behavioral and neuroimaging studies of normal and brain-damaged people, Moscovitch et al., 2005
- Hippocampal PET activations of memory encoding and retrieval: The HIPER model, Lepage et al., 1998
- Event-related fMRI studies of episodic encoding and retrieval: Meta-analyses using activation likelihood estimation, Spaniol et al., 2009
- Reversible information flow across the medial temporal lobe: The hippocampus links cortical modules during memory retrieval, Staresina et al., 2013
- A cortical-hippocampal system for declarative memory, Eichenbaum, 2000
- Components of episodic memory: The contribution of recollection and familiarity, Yonelinas, 2001
- Recollection and familiarity: Examining controversial assumptions and new directions, Yonelinas et al., 2010
- The medial temporal lobe and recognition memory, Eichenbaum et al., 2007
- Remembering the past and imagining the future: Common and distinct neural substrates during event construction and elaboration, Addis et al., 2007
- A New measure of imagination ability: Anatomical brain imaging correlates, Jung et al., 2016
- On the nature of medial temporal lobe contributions to the constructive simulation of future events, Schacter and Addis, 2009
- Creative thought as blind-variation and selective-retention: Combinatorial models of exceptional creativity Simmonton, 2010
- Immortal Truth: A Neuroscientific Critique of Postmodernism, Striegel, 2021
- The Green Eggs and Ham Hypothesis: How Constraints Facilitate Creativity, Haught-Tromp, 2017
- Constraints, Competency and Creativity in the Classroom, Haught-Tromp and Stokes, 2017
- The Scales of the Mind, Striegel, 2021








